| Italian language |
|
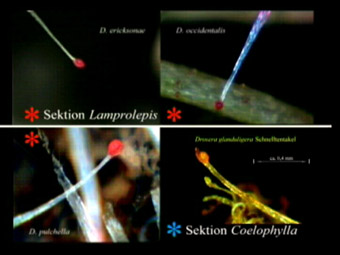
Clandestine Diversity: Snap-Tentacles of the Genus Drosera Hartmeyer, I. & Hartmeyer, S. (2006) CARNIFLORA AUSTRALIS (AUSCPS) 2006/7:4-18 In 1994 we filmed Drosera burmannii in our greenhouse to record the rapid movement of its elongated marginal tentacles for a new video (1). A well known phenomenon even 12 years ago, reported to be similar to Drosera sessilifoliaThelocalyx). After becoming aware of those snap-tentacles without glue-production we surprisingly found some more sundews standing just next to them, actually less eye catching but showing the same tactic, namely the hybrids D. capensis x aliciae and D. rotundifolia x spatulata (synonym: D. tokaiensis) as well as some Pygmy Drosera species (section Lamprolepis). However, fully occupied with our movie we paid no further attention to it, at least until we received a VHS video by Richard Davion in 2003, which brought the rapid tentacles back to mind. Now it was D. glanduligera (section Coelophylla), a minute species and quite difficult to cultivate, which grows naturally near Richard’s property in Adelaide (South Australia). For almost 25 years he tried to convince other CP enthusiasts that the elongated tentacles (which he calls "ribbon-tentacles") of this species need only fractional parts of a second for snapping. 1995 and 99 he even published articles on the subject in "Flytrap News" (2-3) mentioning also the movement of D. burmannii, D. pygmaea and D. callistos. This proved to be in vain, because obviously other CP enthusiasts either ignored or did not believe him – until we showed his VHS recordings on our DVD "A Hunting Veggies Cocktail" (4). Having also been successful in cultivating the minute plants during the following winter season, we started to investigate the phenomenon with a macro-lens and our USB-microscopes. Finally we thoroughly confirmed Richard’s discovery completely, writing an article for the German CPS (GFP) newsletter DAS TAUBLATT (5).
Taking another detailed look at the Drosera species in our collection became a reason to contact some old CP friends. Anja and Holger Hennern showed us Pygmy Drosera photos from their last Australia tour and indeed several more plants showed elongated marginal tentacles without glue production. Photos in literature (i.e.: Plants of Prey (6) or Carnivorous Plants of Australia Vol. 2 (7)) either don’t show them at all, or just coincidentally, and drawings incorrectly show all these tentacles with a glue drop. Obviously the authors did not pay any attention to the phenomenon because there are no further comments. At least the presently most scientific book by the authors Juniper, Robins and Joël „The Carnivorous Plants" (8) mentions – more generally speaking – that several species exist which develop rapid moving, glue-less marginal tentacles e.g. D. burmannii, putting them into one context concerning the evolution from Drosera tentacles to the snap trap of Dionaea. Very interesting! So we started to record plants out of different Drosera sections with macro- and microscope images and spent a lot of time filming real time and time-lapse videos. Then we called on a well known expert for systematics (and an editor of Carnivorous Plant Newsletter - CPN), Dr Jan Schlauer, to ask him for his opinion on the phenomenon as well as for a fitting designation for the obviously mostly overlooked snap tentacles, which he also knew on principle. Somehow surprised about the extent of our observations (which he called "somehow bizarre") he considered that due to their behaviour they could be called catapult- or jerk-tentacles (and in German: Katapult- or Schnelltentakel). We immediately agreed to the German words (especially "Schnelltentakel" is charming) and found also the English names were fitting. Actually after another communication with Richard Davion we decided to keep on using the word snap-tentacles for our English translations. Incidentally some Drosera species show also a rapid movement with glue producing (sometimes elongated) tentacles but these are explicitly not meant. However, sometimes there is indeed a relation between them, as you will see below. Looking at the snap-tentacle behaviour of different species actually not only the Australian Pygmy Drosera species, but nearly all checked "snappers" showed a similar ability to move about as fast as D. burmannii and D.sessilifolia. The 100-fold faster speed of D. glanduligera remains a record and is actually still unique for the genus. Considering the difference between species with or without marginal snap-tentacles – both can be closely related and listed within the same section – leads to an interesting result. Holger Hennern supposed (personal communication) that only basal rosettes develop snap-tentacles, but not stem building species or such with upright pointing trapping leaves because that makes sense for the trapping. In association with flying prey glue is much more effective than a snap-tentacle, which indeed is quite ineffective from the point of view of winged insects. The elongated marginal glue-tentacles of D. binata, D. scorpioides or D. indica are good examples for this. With regard to small Arthropods crawling directly on the soil it’s another story all together. Here the downwards pointing snap-tentacles appear like a set of bars (see photos) that start moving up within seconds when touched by prey, forwarding it directly to the gluey lamina. Once triggered they are either set upright around the leaf margin and complicate escaping the glue, or they bend in like i.e. D. burmannii - and especially effective D. glanduligera – holding on to the prey until it’s really tied up and pressed into the trapping and digesting mucus. In addition this is also a clever method to avoid prey theft by ants, which is a common problem for many Drosera.
Exactly that hypothesis seems to be confirmed by our
investigations.
Considering the shape of a plant (basal rosette or not) for the sections Drosera
and Lamprolepis (see table), it is thus possible to predict
relatively accurately whether a species develops snap-tentacles or not.
Especially interesting in this respect is section Lamprolepis
because also the upright growing species like (conspicuously) D.
scorpioides show precisely at that marginal place elongated
glue-tentacles - also able to move quickly - where i.e. the basal rosettes
D. ericksonae or D. occidentalis develop their
snap-tentacles. Obviously dependent on plant shape we find "snappers"
and "only mucous" plants which are very closely related,
sometimes sharing even the same growing sites. That needs some more
consideration.
The theory of evolution tells us that D. arcturi and D. regia (monotypical sections) are the most archaic Drosera. Neither have marginal snap-tentacles (not yet ?), only uniform "normal" glue-tentacles. The plants in section Ergaleium (bulbous Drosera), Lasiocephala (D. petiolaris and relatives) and Prolifera (Queensland sundews) show the same behaviour. Indeed we find basal rosettes within these sections, but none of them developed snap-tentacles, neither D. schizandra, nor D. whittakeri or D. falconeri. Indeed "snappers" are mainly limited to the Southern Hemisphere. The most northerly reaching species are D. spatulata in the east (Australia to Japan) and D. capillaris in the west (both Americas). Looking at our table below we see that snap-tentacles can be found in Asia, Australia, both Americas and in South Africa. Actually the simultaneous spreading of closely related genera in Australia, South America and Southern Africa (i.e.: the family Proteaceae) is today well known in Biology. The reason are common ancestors some 100 Million years ago, when all the continents (including India) were united as Gondwana. Unfortunately is Antarctica - which also participated in that botanical hotspot - meanwhile too cold for insects and carnivorous plants. However, divided by the following continental shift the surviving genera continued their evolutionary development independently on different continents.
This being considered, one gains the impression of a "modern" Sundew species which, in the days of the Dinosaurs in Gondwana, still developed additional optimised marginal tentacles to effectively catch crawling prey from the soil. The fixation of prey is also a smart way to avoid theft by insects like ants, which also started their successful world-wide development some 100 Million years ago. Maybe there is even causality. In addition the variable behaviour of section Lamprolepis (approx. 40 species) clearly shows an ability to switch - if a new species appears by adaptation – the genes between the production of marginal glue- or snap-tentacles. The important triggers for that amazing adaptation mechanism are environment and prey (mainly if crawling or flying) and thus logically the growing shape of the species. One more observation: if cross-breeding the basal rosette D.aliciae with the upright growing D. capensis the resulting (natural) hybrid D. capensis x aliciae produces rapid snap-tentacles despite development of a stem (within many years). D. binata seedlings show snap-tentacles, but only until the leaves start getting forked. Here is still plenty of room for further investigations. All in all this could be of interest for systematics too and we are looking forward to the discussions on this topic because, last but not least, our USB-microscopes show that the shapes of the investigated snap-tentacle heads are also pleasantly different (see below). From this point of view the trigger mechanism of D. glanduligera looks for example like an evolutionary improvement of the hemisphere-shaped snap-tentacle heads which are typical for the Pygmy Drosera species (relatively similar also to the Southern Australian D. hamiltonii). That structure changed obviously into a stalked ball, which lies moveably on the three-fold subdivided tentacle head, acting as a highly sensitive contact-sensor. Combined with a kind of articulation connecting the now divided tentacle stalk, the movement becomes approximately 100 times faster than in all other species of the genus, which is a really new dimension. Like the authors of "The Carnivorous Plants" (8) state for D. burmannii, we consider also D. glanduligera to be a remarkable development of nature with respect to the development of rapid snap traps in Aldrovanda and Dionaea.
Actually there still remains the question of the trigger of movement in the other snap-tentacles. Tests with a needle show that a trigger mechanism must exist on the tentacle head or the cells connecting head and stalk, but how does it work? With the exception of D. glanduligera all snap-tentacle heads consist of a kind of scoop (lower side mostly flat), with a shape ranging from round to lengthy dependent on the species. On the upper side we find an embossed pillow-like cell-structure (also with various shapes depending on the species) which is in all cases inflated (we call it "jelly-pillow") to varying extents depending on the condition of the plant and the age of the trapping leaf. If that inflated surface is touched, no matter whether by insect or by needle, a shockwave probably moves through the "pillow" and hits the bottom cells, which trigger an action potential that moves on into the tentacle stalk. Or the action potential is produced at the junction between head and stalk, as observed with the mucus producing tentacles. Actually the action potential moves into the stalk, where it causes some particular cell-groups to increase their inner pressure. The whole tentacle starts bending now due to this rapid cell growth (turgor-growth). The development of such action potentials by contact (also by chemical stimulation) has been proved by Prof. Stephen Williams (USA) in the early 70’s with "normal" Drosera tentacles (10-14). Using minute electrodes he measured the action potentials, their movement through the cell tissue and their amazing ability to cause particular remote cells to adapt their pressure, all without the existence of nerves. Therefore it’s very probable that the principles of triggering and movement are very similar for the snap-tentacles which are in fact modified glue-tentacles.
More about snap-tentacles from different continents under our microscopes, magnifications on video in real time and time-lapse, and even more can be found on our new DVD "Drosera – Snap-Tentacles and Runway Lights (German: Drosera – Schnelltentakel und Landescheinwerfer) (9). There we also explain our theory of the function of the yellow emergences of D. hartmeyerorum which also developed from normal glue-tentacles. Last but not least, the trigger-hairs responsible for the snap-trap movement of the Waterwheel Plant and the Venus Flytrap developed from Drosera tentacles too. Another fact that Stephen Williams describes very convincingly by comparative investigations of the family Droseraceae (10-14). The sketches below showing the different shapes of snap-tentacle heads are only thought to highlight the diversity of those structures and we count on your forgiveness for Siggi’s drawing talent. For the support of our investigations and this resulting article we would like to express our special thanks to Richard Davion (who discovererd the fastest tentacle movement in the genus Drosera) and Stephen Williams for their important information and material on the topic. Thanks also to Anja und Holger Hennern who shared their rich "Pygmy experience", and to Jan Schlauer for his opinion and name proposals. We are also grateful to Dr. Eberhard König, who gratis donated some missing species for our investigation, as well as Dr. Barry A. Rice for his support concerning D. capillaris. Last but not least we say thank you so much to Elvis Pöhlmann who spent much time for making the high resolution photos of Drosera species in our CP collection and to Paul Smith for proof reading my poor English.
Our DVD provides even more information than this article. For the first time we describe the reflecting tentacles (runway lights) of Drosera hartmeyerorum with amazing pictures. This are all screenshots from our new film. List of investigated adult plants for our DVD until 2005 New investigations of meanwhile more than 100 species (after our DVD has been released) show that the seedlings of all (investigated) species of section Drosera develop snap-tentacles, not only D. binata. Also the seedlings of section Lasiocephala, where no adult plant possess them. Only the archaic species D. arcturi, D. regia, and strange enough the modern tuberous Drosera do not. D. glanduligera is again special and starts only with glue-tentacles, then intermediate tentacle-heads are developed and only the 3rd or 4th leaf generation shows the ready developed spring trap. An upgrading article on this topic is ready written and only waiting to be published. The story goes on.
Photos: Elvis Pöhlmann & S. Hartmeyer – Tables and sketches: S. Hartmeyer Copyright (2005) I. and S. Hartmeyer Bibliography: Hartmeyer, I., Hartmeyer, S., (1995) Reiseziel Insektivoren (Englisch: Beautiful and Hungry – Part 2), private DVD Davion, R., (1995) Now you see it - Now you don’t, FLYTRAP NEWS (CPSNSW) Vol. 8/4:17 Davion, R., (1999) That damned elusive Pimpernel, FLYTRAP NEWS (CPSNSW) Vol. 13/1: 10 Hartmeyer, I., Hartmeyer, S., (2004) Quer durch’s Karnivorenbeet" (Englisch: A HUNTING VEGGIES Cocktail), private DVD Hartmeyer, I. & Hartmeyer, S., (2005) Drosera glanduligera: Der Sonnentau mit "Schnapp-Tentakeln", DAS TAUBLATT (GFP) 2005/2: 34-38 Erickson, R., (1968) Plants of Prey, University of Western Australia Press Lowrie, A. (1989) Carnivorous Plants of Australia Vol. 2, University of Western Australia Press B.E. Juniper, R.J. Robins. D.M. Joel, (1989) The Carnivorous Plants, Academic Press London: 303 Hartmeyer, I. & Hartmeyer, S., (2006) Drosera - Schnelltentakel und Landescheinwerfer" (Englisch: Drosera – Snap-Tentacles and Runway-Lights), private DVD Hartmeyer, I. & Hartmeyer, S., (2002) The International Carnivorous Plant Conference – 2002 in Tokyo, private DVD Williams, S.E., Pickard, B.G., Connections and Barriers between Cells of Drosera Tentacles in Relation to Their Electrophysiology, Planta (Berl.) 116, 1-16 (1974) Williams, S.E., Comparative Sensory Physiology of the Droseraceae - The Evolution of a Plant Sensory System, Proceedings of the American Philosophical Society, Vol. 120 No. 3, June 1976 Williams, S.E., Pickard, B.G., The Role of Action Potentials in the Control of Capture Movements of Drosera and Dionaea, Plant Growth Substances 1979, Springer Verlag Berlin-Heidelberg-New York Williams, S.E., Albert, V.A., Chase, M.W., Relationships of Droseraceae: A Cladistic Analysis of RBCL Sequence and Morphological Data, American Journal of Botany, 81(8): 1027-1037, 1994 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||