| Schnelltentakel & Mikroskop | English language | Italian language | Artikel und Berichte |
|
|
Verborgene
Vielfalt: Die Schnelltentakel der Gattung
Drosera Alle Bilder stammen von unserer DVD "DROSERA - Schnelltentakel und Landescheinwerfer" Als wir 1994 in unserem Gewächshaus Drosera burmannii
filmten, um die schnelle Bewegung der auffällig über die Blattfläche
verlängerten Tentakel zu dokumentieren (1), war dies ein
bekanntes Phänomen, das ähnlich auch von Drosera sessilifolia
(beide Sektion Thelocalyx) bekannt war. Sensibilisiert auf diese
Tentakel ohne Fangschleim fanden wir solche - wenn auch weniger
ausgeprägt - zu unserer Überraschung auch bei anderen Sonnentau, die
zufällig in Töpfen daneben standen, nämlich den Hybriden D. capensis x aliciae
und D. rotundifolia x spatulata
(auch D. tokaiensis), sowie bei einigen Zwergsonnentau (Sektion Lamprolepis).
Da wir jedoch mit unserem Filmprojekt voll beschäftigt waren, maßen wir
dem Phänomen keine weitere Bedeutung bei, jedenfalls bis uns ein
VHS-Video von Richard Davion im Jahr 2003 die schnellen Tentakel
schlagartig wieder ins Bewusstsein brachte. Jetzt war es D.
glanduligera (Sektion Coelophylla), eine winzige, schwer zu
kultivierende Art, die gleich neben Richards Grundstück in Adelaide
(Südaustralien) wächst. Er hatte fast 25 Jahre lang versucht, anderen
Karnivorenkennern zu vermitteln, dass die verlängerten Tentakel dieser
Art (er nennt sie im Englischen „Ribbon-Tentacles") sogar im
Sekundenbruchteil vorklappen und schrieb bereits 1995 und 99 Artikel
darüber in der „Flytrap News" (2-3), worin er neben D.
burmannii auch bereits D. pygmaea und D. callistos
erwähnt. Vergeblich, denn es glaubte wohl niemand recht daran, bis wir
seine Aufnahmen auf unserer DVD „Quer durch’s Karnivorenbeet" (4)
zeigten und das Phänomen mit Makro und USB-Mikroskop genauer
untersuchten, um es dann in einem Artikel für das TAUBLATT (5)
der GFP voll und ganz bestätigen zu können.
Natürlich schauten wir jetzt nochmals genauer auf die Drosera in unserer Sammlung und was wir da fanden veranlasste uns, einige alte Freunde zu kontaktieren. Mit Anja und Holger Hennern hatten wir bereits über ihre australischen Standortfotos gesprochen. Darauf zeigen einige Zwergsonnentau ebenfalls marginale Langtentakel ohne Klebstoffproduktion. Fotos in Sachbüchern über die „Pygmys" (z.B.: Plants of Prey (6) oder Carnivorous Plants of Australia Vol. 2 (7)) zeigen diese nur zufällig und unkommentiert, Zeichnungen stellen alle inkorrekt als Klebstofftentakel dar. Diese Autoren schenkten dem Phänomen keine Beachtung, denn es gibt auch keinerlei weitere Bemerkungen dazu. Im derzeit wohl wissenschaftlichsten Buch der Autoren Juniper, Robins und Joël „The Carnivorous Plants" (8) findet sich immerhin ein recht allgemein gehaltener Hinweis auf die leimlosen, schnell beweglichen Randtentakel diverser Arten, mit D. burmannii als Beispiel. Deren Eigenschaften werden dort als mögliche Zwischenstation bei der Evolution von den Leimfallen des Sonnentau zur Klappfalle der Venus Fliegenfalle gesehen. Sehr interessant! Also begannen wir, das Phänomen in unserer Sammlung mit Makro- und Mikroskopaufnahmen zu dokumentieren. Als nächstes riefen wir den bekannten Systematiker (und ein Editor des Carnivorous Plant Newsletter) Dr. Jan Schlauer an, da uns seine Meinung zum Thema interessierte und wir noch eine passende Bezeichnung für die offensichtlich häufig übersehenen „Schnapptentakel" suchten, die ihm prinzipiell natürlich bekannt waren. Etwas überrascht über das Ausmaß unserer - in seinen Augen „etwas skurrilen" - Beobachtungen lauteten seine Vorschläge: Katapult- oder Schnelltentakel (für schnell, aber auch für empor schnellen), was uns beides auf Anhieb passend erschien und daher ab jetzt von uns verwendet wird. Übrigens können sich bei einigen Sonnentau die äußeren Klebstoff produzierenden Tentakel ebenfalls schnell und deutlich sichtbar bewegen. Diese sind hier jedoch ausdrücklich nicht gemeint, obwohl es auch da bei einigen Arten einen engen Zusammenhang gibt, wie wir gleich zeigen werden.
Mit der Videokamera filmten wir die Bewegung der Schnelltentakel bei verschiedenen Arten. Tatsächlich waren dabei nicht nur die australischen Zwergsonnentau, sondern fast alle getesteten Arten zu ähnlich schnellen Bewegungen fähig wie D. burmannii und D.sessilifolia. Die 100-fach schnellere Geschwindigkeit von D. glanduligera bleibt aber nach wie vor einzigartig in der Gattung. Zu einem interessanten Ergebnis führt die Auswertung der Daten, basierend auf der Frage, worin sich Arten mit Katapulttentakeln von solchen mit „nur" Klebstofftentakeln unterscheiden, wobei Vertreter beider Varianten durchaus der gleichen Sektion zugeordnet sein können. Dazu fanden wir gar keine Literatur. Holger Hennern hatte uns gegenüber bereits die Vermutung geäußert, dass nur am Boden aufliegende Rosetten Schnelltentakel ausbilden, nicht aber Stamm bildende Arten oder solche mit gestielten, aufrecht stehenden Blättern. Das macht für den Fangvorgang nämlich durchaus Sinn: bei Berührung im Flug sind Insekten mit Leim wesentlich effektiver zu fangen als mit Schnelltentakeln, die aus der Sicht von Fluginsekten immer noch ziemlich lahm sind. Die langen Klebstofftentakel an den Blattenden von D. binata, D. scorpioides oder D. indica sind hierfür ein gutes Beispiel. Bei am Boden krabbelnden Kleininsekten sieht das allerdings ganz anders aus. Hier erzeugen die nach unten geklappten Schnelltentakel teils ein regelrechtes Sperrgitter, welches bei Berührung in einigen Sekunden nach oben „schnellt", um die Beute zum Klebstoff zu transportieren. Einmal ausgelöst werden sie entweder über den Blattrand aufgerichtet und erschweren zusätzlich ein Entkommen aus dem Leim, oder sie biegen sich weiter, wie bei D. burmannii - und besonders effektiv bei D. glanduligera - wobei sie die Beute effektiv festhalten und regelrecht in den Fangschleim pressen.
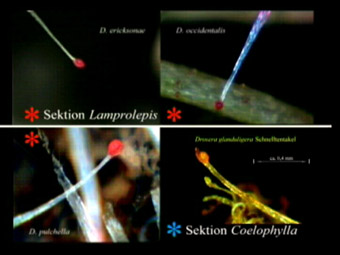
Und genau dies bestätigen jetzt unsere Aufnahmen. So lässt sich zum Beispiel recht verlässlich in den Sektionen Drosera und Lamprolepis (siehe Tabelle) vorhersagen, ob eine Art Schnelltentakel ausbildet oder nicht, wenn man die Wuchsform berücksichtigt. Interessant ist in diesem Zusammenhang besonders die Sektion Lamprolepis, denn hier zeigen auch die aufrecht wachsenden Arten wie (besonders auffällig) D. scorpioides genau dort, wo z.B. bei der Bodenrosette D. ericksonae am Blatt die Schnelltentakel sitzen, stark verlängerte marginale Tentakel, welche ebenfalls relativ schnell beweglich sind, allerdings produzieren diese Klebstoff. Die Ausbildung beider Tentakelformen tritt hier also offensichtlich bei sehr nah verwandten Arten, mitunter an gemeinsamen Standorten, je nach Wuchsform auf. Die uns bekannten abstammungsgeschichtlich ältesten Arten D. arcturi und D. regia (monotypische Sektionen) besitzen (noch?) keine typischen Marginaltentakel, sondern recht gleichförmige „normale" Leimtentakel. Ebenso alle Vertreter der Sektionen Ergaleium (Knollen bildende), Lasiocephala (D. petiolaris und Verwandte) und Prolifera (Nord-Queensland Drosera). Allen sind auch Bodenrosetten zugeordnet, aber keine davon, weder D. schizandra, noch D. whittakeri oder D. falconeri bildet Schnelltentakel aus. Überhaupt sind diese im Wesentlichen auf Arten in der südlichen Hemisphäre begrenzt. Im Osten ist D. spatulata (Australien bis Japan) und im Westen D. capillaris (beide Amerika) die am weitesten in den Norden vordringende Art. Wie unsere unten stehende Tabelle zeigt, finden sich Schnelltentakel in Asien, Australien, beiden Amerika und in Südafrika. Die gleichzeitige Verbreitung eng verwandter Gattungen in Australien, Südamerika und Südafrika (wie z.B. die Familie Proteaceae) ist in der Biologie schon lange bekannt. Das ist auf gemeinsame Vorfahren aus Zeiten vor etwa 100 Millionen Jahren zurückzuführen, als die genannten Kontinente inklusive Indien noch in Gondwana vereinigt waren. Leider ist es in der Antarktis, die ebenfalls an diesem biologischen „Hotspot" beteiligt war, inzwischen zu kalt für Insekten und Karnivoren. Getrennt durch die Kontinentalverschiebung haben sich die Arten dann durch evolutionäre Prozesse unabhängig von einander weiter entwickelt.
Es sieht also ganz danach aus, als hätten einige „modernere" Sonnentau noch in Gondwana, zur Zeit der Dinosaurier, die optimierten Marginaltentakel „erfunden" und die Fähigkeit zu deren Ausbildung als Schnell- oder Leimtentakel im Genom gespeichert. In diese Zeit fällt auch die erfolgreiche Ausbreitung der Ameisen, möglicherweise stehen diese Entwicklungen in Zusammenhang. Die Betrachtung der Sektion Lamprolepis legt nahe, dass die Nachkommen je nach Umgebungsbedingungen und Beute (Bodeninsekten oder Fluginsekten) offensichtlich in der Lage sind, die jeweils für die günstigere Wuchsform benötigten Gene ein- oder auszuschalten, sobald durch Anpassung eine neue Art entsteht. Dazu noch eine Beobachtung: bei Kreuzungen aus Bodenrosette (z.B. D. aliciae) und aufrechter Pflanze (z.B. D. capensis) werden die Schnelltentakel übrigens vererbt, selbst wenn die entstandene Kreuzung Stämmchen bildet, wie in diesem Beispiel die Naturhybride D. capensis x aliciae. Bei D. binata bilden nur die Sämlinge Schnelltentakel, bis sich die Blätter zu gabeln beginnen. Hier ist aber noch viel Raum für weitergehende Forschungen.
Dies alles ist sicher nicht ganz uninteressant für die Systematiker unter uns und wir freuen uns schon auf die Diskussionen zu diesem Thema, denn „last but not least" auch die Form der Schnelltentakel-Köpfe ist erfreulich unterschiedlich (siehe unten), wie Untersuchungen mit unseren USB-Mikroskopen zeigen. So scheint z.B. der Triggermechanismus von D. glanduligera aus dieser Sicht eine Weiterentwicklung der für die Zwergsonnentau typischen Halbkugel-Struktur des Tentakelkopfes zu sein (die ähnlich auch die südaustralische D. hamiltonii zeigt). Diese entwickelte sich zu einer jetzt gestielten Kugel, welche beweglich auf der dreifach unterteilten Tentakelspitze aufliegt und bei der geringsten Bewegung als Sensor und Impulsgeber funktioniert. In Verbindung mit einer Art Gelenk im Tentakelstiel wird die Bewegung um etwa den Faktor 100 schneller, als bei allen anderen Vertretern der Gattung, ein echter Quantensprung. Wie die Autoren von „The Carnivorous Plants" (8) bereits bei D. burmannii bemerkten, sehen wir auch bei D. glanduligera einen bemerkenswerten Fingerzeig der Natur auf die Entwicklung der Klappfallen bei Aldrovanda und Dionaea. Es bleibt die Frage, wodurch die Bewegung der anderen Schnelltentakel ausgelöst wird. Versuche mit einer Nadel zeigen, dass sich ein Triggermechanismus im oder am Tentakelkopf befindet, aber wie funktioniert er? Abgesehen von D. glanduligera bestehen alle Köpfe der Schnelltentakel aus einer Art Schaufel (Unterseite meist flach), deren Form je nach Spezies von rund bis langgestreckt variiert. Darauf befindet sich eine erhabene, kissenförmige Zellstruktur, deren Form zwar ebenfalls je nach Art variiert, welche jedoch immer die Eigenschaften eines Schwellkörpers zeigt, denn je nach Zustand der Pflanze kann diese (von uns auch als „Gelkissen" bezeichnet) unterschiedlich stark aufgebläht sein. Bei Berührung der Oberfläche, egal ob durch Insekt oder Nadel, entsteht möglicherweise eine Druckwelle, die dann in den unteren Zellen einen elektrischen Impuls auslöst, welcher in den Tentakelstiel geleitet wird. Oder die Bewegung des ganzen Kopfes löst (wie bei den Leimtentakeln) an der Verbindung zum Tentakelstiel einen Impuls aus. Jedenfalls setzt sich dieser fort, wodurch an bestimmten Stellen im Tentakelstiel Zellgruppen veranlasst werden, ihren Innendruck zu verändern. Der Tentakel biegt sich jetzt spontan durch diese Art des Zellwachstums. Elektrische Potenziale nach Berührung (oder auch bei chemischer Reizung) wurden bereits Anfang der 70er Jahre von Prof. Stephen Williams (USA) an normalen Leimtentakeln nachgewiesen (10-14). Mit winzigen Elektroden hat er die Stromimpulse und deren Weiterleitung im Zellgewebe gemessen, sowie deren Eigenschaft, bestimmte räumlich entfernte Zellen zur Druckänderung zu veranlassen. Und alles ganz ohne Nervenbahnen! Es ist daher sehr wahrscheinlich, dass dieses Prinzip auch für die Bewegung der - lediglich modifizierten - Schnelltentakel gilt.
Weitere Mikroskopvergleiche der Tentakelköpfe von verschiedenen Kontinenten, sowie Makroaufnahmen dazu in Echtzeit und Zeitraffer gibt es auf unserer DVD „Drosera - Schnelltentakel und Landescheinwerfer" (Englisch: Drosera – Snap-Tentacles and Runway-Lights) (9) zu sehen. Außerdem erläutern wir darauf unsere Theorie zur Funktion der gelben Emergenzen von D. hartmeyerorum, die sich ja ebenfalls aus normalen Leimtentakeln entwickelt haben. Übrigens, auch die Triggerhaare, welche die Klappbewegung der Wasserfalle und der Venus Fliegenfalle auslösen, haben sich aus den Tentakeln des Sonnentau entwickelt, wie Stephen Williams bei seinen vergleichenden Forschungen (10-14) überzeugend darstellt. Die unten stehenden Zeichnungen der verschiedenen von uns gefundenen Tentakelformen sollen lediglich die groben Unterschiede deutlich machen, weshalb wir um Nachsicht bitten, was das zeichnerische Talent angeht. Wir sehen noch viel Raum für weitergehende Forschungen zum Thema und hoffen, einen Anreiz dazu geschaffen zu haben. Tatsächlich erscheint es uns immer noch unfassbar, dass sowohl die erstaunliche Vielfalt, als auch die spezielle Struktur der Schnelltentakel in der Fachliteratur quasi keine Rolle spielt und deren Vorhandensein in den meisten Artbeschreibungen schlicht fehlt.
Anmerkung zum Thema Landescheinwerfer Unsere DVD bietet noch mehr
als in diesem Artikel zu lesen ist. Dazu gehört auch die Beschreibung der
"Landescheinwerfer" von Drosera hartmeyerorum mit
verblüffenden Bildern. Alle Aufnahmen sind Screenshots aus unserem neuen
Film. Zu diesem Phänomen gibt es - außer auf unserer DVD bisher weder
Literatur noch Bilder! Von uns bis 2005 für die DVD untersuchte ausgewachsene Pflanzen nach Sektionen geordnet Neue Resultate von inzwischen mehr als
100 Arten (nach der Veröffentlichung unserer DVD) zeigen, dass alle
untersuchten Sämlinge der Sektion Drosera Schnelltentakel
entwickeln, also nicht nur D. binata. Auch die Sämlinge der
Sektion Lasiocephala, deren ausgewachsene Arten keinerlei
drüsenlose Marginaltentakel bilden. Nur die archaischen Arten D. arcturi, D. regia,
die Sektion Prolifera und seltsamerweise die modernen
Knollendrosera machen das nicht. D.
glanduligera ist wiederum eine Ausnahme und bildet erst marginale
Leimtentakel, dann Übergangsformen und erst die dritte oder vierte
Blattgeneration bildet fertige Schnapptentakel aus. Ein neuer Artikel der
alles auf den neuesten Stand bring ist bereits geschrieben und wartet auf
seine Veröffentlichung. Die Geschichte bleibt spannend!
Digitalfotos zum Artikel: Elvis
Pöhlmann – Videobilder: Siegfried Hartmeyer Literatur: Hartmeyer, I., Hartmeyer, S., (1995) Reiseziel Insektivoren (Englisch: Beautiful and Hungry – Part 2), private DVD Davion, R., (1995) Now you see it - Now you don’t, FLYTRAP NEWS (CPSNSW) Vol. 8/4:17 Davion, R., (1999) That damned elusive Pimpernel, FLYTRAP NEWS (CPSNSW) Vol. 13/1:10 Hartmeyer, I., Hartmeyer, S., (2004) Quer durch’s Karnivorenbeet" (Englisch: A HUNTING VEGGIES Cocktail), private DVD Hartmeyer, I. & Hartmeyer, S., (2005) Drosera glanduligera: Der Sonnentau mit "Schnapp-Tentakeln", DAS TAUBLATT (GFP) 2005/2: 34-38 Erickson, R., (1968) Plants of Prey, University of Western Australia Press Lowrie, A. (1989) Carnivorous Plants of Australia Vol. 2, University of Western Australia Press B.E. Juniper, R.J. Robins. D.M. Joel, (1989) The Carnivorous Plants, Academic Press London: 303 Hartmeyer, I. & Hartmeyer, S., (2006) Drosera - Schnelltentakel und Landescheinwerfer" (Englisch: Drosera – Snap-Tentacles and Runway-Lights), private DVD Hartmeyer, I. & Hartmeyer, S., (2002) The International Carnivorous Plant Conference – 2002 in Tokyo - Droseraceae Symposium, private DVD Williams, S.E., Pickard, B.G., Connections and Barriers between Cells of Drosera Tentacles in Relation to Their Electrophysiology, Planta (Berl.) 116, 1-16 (1974) Williams, S.E., Comparative Sensory Physiology of the Droseraceae - The Evolution of a Plant Sensory System, Proceedings of the American Philosophical Society, Vol. 120 No. 3, June 1976 Williams, S.E., Pickard, B.G., The Role of Action Potentials in the Control of Capture Movements of Drosera and Dionaea, Plant Growth Substances 1979, Springer Verlag Berlin-Heidelberg-New York Williams, S.E., Albert, V.A., Chase,
M.W., Relationships of Droseraceae: A Cladistic Analysis of RBCL
Sequence and Morphological Data, American Journal of Botany, 81(8):
1027-1037, 1994
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||